1. Le Neurone
Le neurone est le messager de l'influx nerveux, il se compose d'un corps cellulaire, appelé soma, d'un axone et de dendrites.
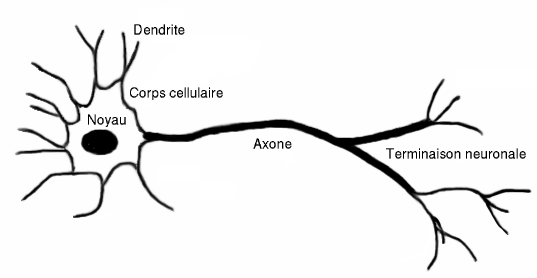
L'axone, se ramifie en terminaisons neuronales, dont l'ensemble est appelé arborisation terminale. Les terminaisons neuronales se terminent par de multiples boutons synaptiques. Les dendrites sont des prolongements du soma des neurones . Deux neurones peuvent se connecter entre eux, au niveau des boutons pré-synaptiques de l'un et des boutons post-synaptiques de l'autre. Ils forment ainsi une synapse. C'est une connection axodendritique. Les dendrites reçoivent des entrées d'autres neurones au niveau des synapses dont elles constituent l'élément post-synaptique.
Le neurone est soumis à un potentiel de membrane, ce potentiel est dû à une répartition inégale des ions (Na+ et K+) de part et d’autre de la membrane. Pour les cellules nerveuses ce potentiel de repos est de l'ordre de -70mV. Le potentiel de repos est principalement dû aux ions K+, car la membrane est beaucoup plus perméable à cet ion qu’aux ions Na+.
Le message nerveux se transmet à partir du segment initial (zone du soma à la base de l'axone) sous forme d'un potentiel électrique nommé : potentiel d'action (PA).
2. Le Potentiel d´Action
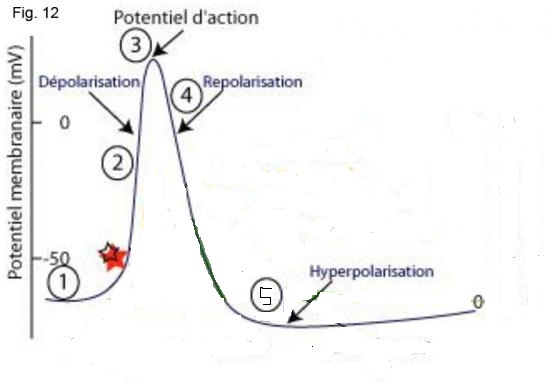
Lorsque le potentiel membranaire (1) franchit un seuil de dépolarisation les canaux Na+ s'ouvrent (étoile rouge).
Sous l'effet du gradient électrochimique, comme la concentration d'ions Na+ à l’extérieur de la cellule est plus forte qu'à l’intérieur, des ions Na+ entrent à l'intérieur. Cet apport de charges positives crée une dépolarisation (une augmentation du potentiel) progressive (2).
Ensuite, les canaux K+ voltage-dépendant s'ouvrent petit à petit peu après les canaux Na+. La concentration de K+ est plus élevée à l’intérieur qu'à l’extérieur de la cellule, donc les ions K+ sortent. Ainsi l’augmentation du potentiel est faiblement contrebalancée jusqu'à ce que les canaux Na+ commencent à s'inactiver ce qui correspond au maximum du potentiel d'action (3). Les canaux K+ sont alors complètement ouvert, la phase de repolarisation commence (4). Cependant le potentiel d'action ne retombe pas à la valeur de potentiel de repos immédiatement. En effet, les canaux K+ sont encore ouvert, c'est la phase d’hyperpolarisation (5). Il faut noter que, comme les canaux Na+ sont inactivés pendant le début de la phase d'hyperpolarisation, c'est a dire qu'ils ne peuvent ni s'ouvrir ni se fermer (et donc entraîner une dépolarisation etc.. ), il est impossible qu'un autre potentiel d'action se superpose à celui-ci, indépendamment de la «force» des entrées: c'est la période réfractaire absolue. De plus, pendant la phase d'hyperpolarisation les canaux K+ étant encore ouvert, la membrane ne peut être stimulée que par des entrées plus fortes pour pouvoir produire un nouveau potentiel d'action, ce qui demande généralement plus de temps et d’énergie, c'est la période réfractaire relative.
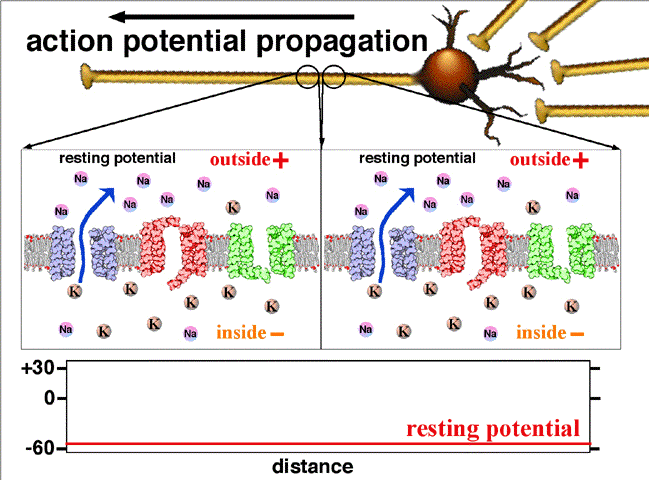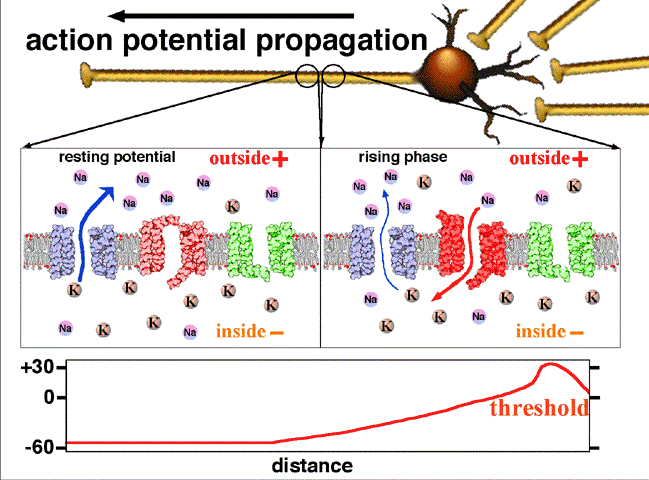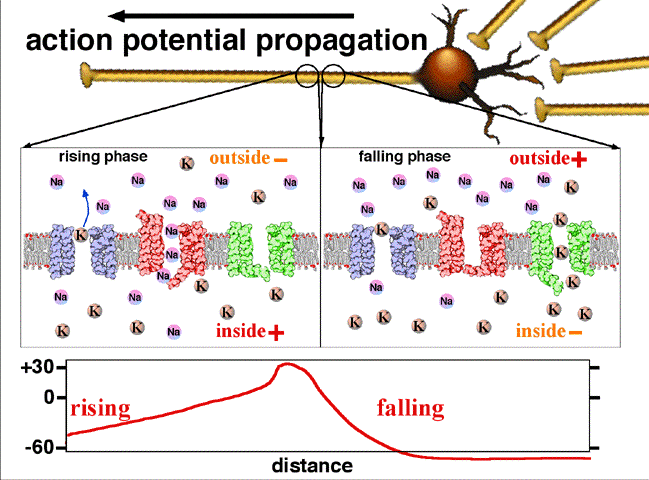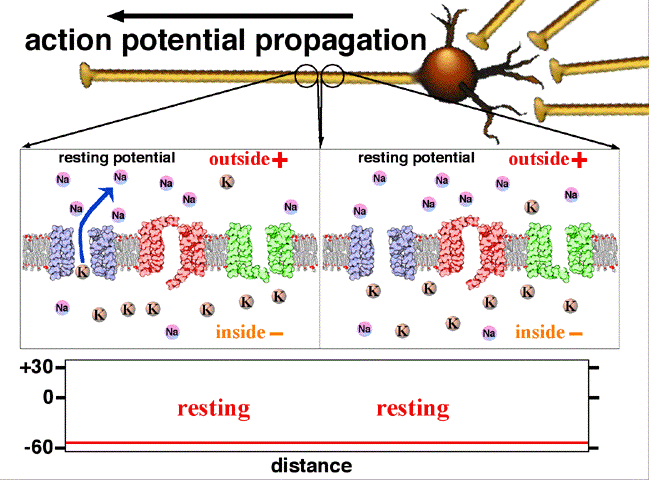

Les charges positives s'éliminent au fur et à mesure de la progression de l'influx. Dans les axones non myélinisés, il existe sur la membrane des pompes voltage-dépendant permettant de régénérer l'intensité d'origine de l'influx nerveux, jusqu'au bout de l'axone.
La période réfractaire assure que le potentiel se déplace à sens unique, vers les terminaisons pré-synaptique, et qu'une période de « repos » est instaurée entre chaque PA. Il faut noter que certains axones peuvent être très longs. Dans ce cas ils sont généralement myélinisés. C'est-à-dire que des gaines régulières de myéline entourent l'axone par endroits. Ces gaines, isolantes, permettent au potentiel d'action de « sauter » d'un espace non myélinisé (nœud de Ranvier) à l'autre. On parle alors de conduction saltatoire. Ainsi le potentiel d'action ne se reforme (via le processus ionique décrit ci-avant) qu'au niveau du nœud de Ranvier suivant. Cela se traduit par un gain de temps (le processus ionique étant relativement lent) et donc une transmission du signal jusqu'à 50 fois plus rapide que pour un axone non myélinisé.
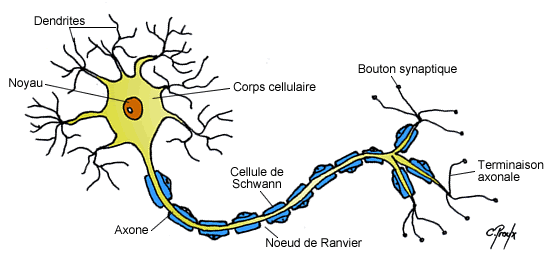

Les charges positives (Sodium) se déplace au bout de l'axone suivant le gradient électrochimique. Les gaines de myélines isolent l'axone, rien ne sort, rien de rentre à leur niveau. Entre deux gaines (noeud de Ranvier), on retrouve des pompes sodiques voltage dépendant régénérant le potentiel d'action initial.
Le potentiel d’action a toujours la même amplitude quel que soit les entrées qui l’ont déclenchées. Cette amplitude correspond au temps nécessaire à la fermeture de la porte d’inactivation pendant lequel les canaux Na+ sont ouverts. Ainsi si le seuil est atteint, ou même dépassé, il y aura un potentiel d’action, mais s’il n’est pas atteint alors il n’y aura rien du tout. On dit pour cela que le potentiel d'action obeit a la loi du tout ou rien.
Après le déclenchement d'un PA le potentiel de membrane retombe progressivement a sa valeur de repos, sous l'action d'une pompe : la pompe NaKAtpase.
3. La Synapse
Le potentiel d'action se propage tout le long de l'axone. Lorsqu'il arrive dans les terminaisons pré-synaptiques, il modifie localement le potentiel de membrane ce qui ouvre des canaux Ca2+ lorsque le seuil d'activation est atteint. Comme la concentration de Ca2+ est plus forte à l'extérieur que dans le bouton pré-synaptique, du Ca2+ rentre dans ce dernier (gradient électrochimique). Le bouton pré-synaptique renferme de nombreuses vésicules synaptiques qui contiennent les neurotransmetteurs. L'entrée du Ca2+ dans le bouton permet aux vésicules de se déplacer et de fusionner avec la membrane. Les neurotransmetteurs sont ainsi libérés par exocytose dans la fente synaptique. Le neurotransmetteur se propage dans la fente synaptique pour aller se fixer à des récepteurs spécifiques sur la membrane post-synaptique (bouton post-synaptique).
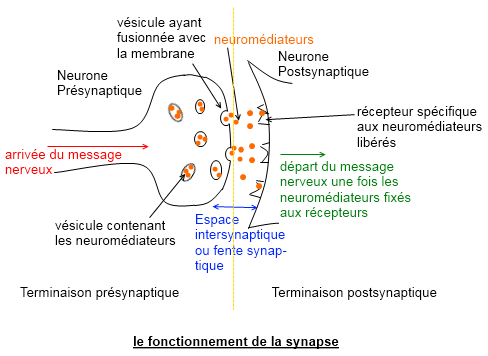
Les récepteurs sont liés à des canaux ioniques. Il en existe deux types, les ionotropiques et les métabotropiques. La transmission du signal d'un bouton pré- à post-synaptique ne se fait pas de manière instantanée. On note ainsi un temps de transmission au niveau de la synapse appelé délai synaptique directement lié au temps que met le neurotransmetteur à traverser la fente synaptique, à son "recyclage" et à la conversion du signal chimique en signal électrique par les récepteurs post-synaptiques. Chaque bouton post-synaptique possède un seul type de récepteur (ionotropique ou bien métabotropique). Ainsi la spécificité d'une terminaison réside dans le couplage d'un neurotransmetteur et d'un récepteur. Par conséquent, un bouton post-synaptique est seulement inhibiteur ou excitateur. Ensuite tout dépend du gradient électrochimique. L'ouverture des canaux permet à l'ion spécifique de rentrer dans le bouton post-synaptique ou bien d'en sortir. Ces déplacements d'ions engendrent des différences locales de potentiels électrochimiques qui provoquent soit une dépolarisation (site excitateur) soit une hyperpolarisation (site inhibiteur) de la membrane post-synaptique, créées selon le sens de déplacement de l'ion et de sa nature. La structure du contact entre deux neurones, pré- et post-synaptique, se compose d'une multitude de synapses du même type: uniquement inhibitrices ou excitatrices avec le même couple de neurotransmetteur et de récepteur pour toutes les synapses. La différence entre chaque synapse réside dans: la quantité de neurotransmetteurs émis, le nombre de canaux ioniques (ie la capacité à absorber ou relâcher les ions), le nettoyage ou/et la récupération des neurotransmetteurs dans la fente synaptique, de la taille de la fente, ainsi que la distance de la synapse au soma du neurone récepteur.
On a vu plus haut, que les dendrites sont la partie post-synaptique de la synapse.
Le potentiel post-synaptique contrairement au potentiel d'action, n’obéit pas à la loi du tout ou rien. Dans les dendrites, le potentiel se propage avec décrément (on parle de potentiels graduels). C'est pour cela que la distance entre la synapse et le soma est importante, car plus la connexion est loin du soma, plus les pertes sont importantes.
C'est ensuite le soma qui intégre les potentiels reçus, et si ils sont suffisants pour élever le potentiel de membrane jusqu'à sa valeur seuil, un potentiel d'action est déclenché au niveau du segment initial.